
 The Early Bird Assembling the Tree of Life research group just published a landmark study in Science looking at the higher-level relationships among birds (Hackett et al. 2008). I think this is the most comprehensive molecular phylogeny of the avian Tree of Life (TOL) to date, and it confirms and validates many new hypotheses of bird inter-relationships that have been proposed in the past ten years.
The Early Bird Assembling the Tree of Life research group just published a landmark study in Science looking at the higher-level relationships among birds (Hackett et al. 2008). I think this is the most comprehensive molecular phylogeny of the avian Tree of Life (TOL) to date, and it confirms and validates many new hypotheses of bird inter-relationships that have been proposed in the past ten years.This work has already hit the blogosphere, with reviews by Grrlscientist, Christopher Taylor, and Greg Laden. I’d like to do two things with this post. First, I’d like to step through all of the major clades of the avian tree. I want to discuss the evidence supporting each clade, including why groupings that seem to clash with previous hypotheses (and common sense) may actually be correct. Second, I have a bad case of SIWOTI syndrome and want to correct some of the popular press reportings on the subject – particularly those that claim this paper is full of ‘new’ results. But I’ll keep it mostly about the birds, don’t worry.
So what new evidence does this paper bring to the table? The avian TOL has been tackled by many studies before. These include several molecular studies using mitochondrial and nuclear DNA, which have been increasing in breadth of sampling and depth of resolution of the tree over the years, including at least three other studies this year (Morgan-Richards et al, Brown et al, Chojnowski et al). Phylogenies based on morphological characters are also still being done, with the most complex recent work I know of being Livezey and Zusi (2007) analyzing a mere 2954 morphological traits.
The Hackett et al. study brings to the table the most comprehensive molecular phylogeny to date. In comparison to the 5007 base pairs of DNA sequence from five nuclear loci in 87 bird species used in Ericson et al. (2006), Hackett et al. (2008) use 32 thousand base pairs from 19 nuclear loci in 171 species. That’s a lot of pipetting! The chief benefit of using so many more loci is resolving power – each locus adds an independent dataset to the tree, strengthening clades and resolving the relationships in the tree. Compare the Ericson tree with the Hackett tree, and you’ll see the difference. The Ericson et al. tree gets many of the basic groupings, but leaves many relationships as polytomies (many branches from a single node on the tree) because they don’t have the data to resolve the true bifurcating tree. The Hackett et al. tree still contains a few polytomies, but the resolution of clades is some the best yet seen.
Ericson et al. 2006

Hackett et al. 2008
Hackett et al. 2008

Hackett et al. (2008) actually present three trees in their work, which you may have seen on other posts. One is the real nuts and bolts tree, the actual results that show every taxa, as well as the support values for each node on the tree.

They then simplify the tree to give the big picture in terms of the explosive radiation of Neoaves, as seen by the very short distances between different groups on the tree, and in terms of the amount of variation in divergence among taxa. The length of each branch in this tree corresponds to the number of substitions in the DNA of that species, or roughly how divergent it is from the node it connects to. The species with really long branch lengths have undergone a lot of change and may be difficult to properly place in the tree.
This explosive radiation is important, because it has prevented us from fully understanding the relationships of the different bird groups. Figuring it out involves a lot of data – we need to find loci that evolve fast enough to ‘capture’ the divergence events in their genealogies, but that do not evolve so fast that relationships become masked by changing and re-changing the sequences (there are only four possibilities at each step in the sequence, after all). Basically, every time a population splits, the DNA in each population begins evolving in different ways at a certain rate. If the two populations split again before the DNA in each has evolved to the point where it is distinct from the DNA in the other lineage, then the history of the first splitting event is not recorded in the sequence data. Rapid radiations produce many such splitting events in a short period of time, making it very difficult to figure out just what went on using only sequence data.
The third tree is the most interesting for studying the taxonomy of birds. This one collapses all of the poorly supported nodes in the main tree, and gives the taxonomic history for each group sampled. This shows how our previous notions of avian taxonomy have been all scrambled, particularly with certain groups such as Pelecaniformes and Gruiformes.

I've combined pieces of these trees where relevant to aid discussion. If any of them are too small to read, just click on any picture to view it full size. The discussion below is only my take on some of these phylogenetic hypotheses. Many of them I have never researched before, and because of this I certainly may have missed key literature. If you think something is wrong - comment on it. Finally, I've included many pictures from the web to look at while reading (all should be properly cited to source). While reading about new phylogenetic hypotheses, take a look at the organisms involved, and compare their similarities and differences in your mind while you read about the evidence supporting them. To aid in this, as I don't expect most people to have a familiarity with every bird family out there, I've linked every family to a web page about them - mostly those from Don Roberson's excellent Bird Families of the World, and where he doesn't cover, Wikipedia.
Starting with the basal branch on the avian TOL:
Paleognathae
The oldest branch on the tree separates Paleognathae from the remaining birds, Neognathae. Paleognathae are the ratites, including the iconic flightless Ostrich, rheas, Emu, cassowaries, and the kiwi. Also in Paleognathae are the tinamou, an odd-looking group of around 50 species of flighted Neotropical birds, with heads too small for their rotund bodies. The separation of Paleognathae from the remaining extant birds has strong consensus, on account of various morphological characters, particularly skeletal characters in the roof of the mouth (gnath refers to jaw, Paleognathae are the old jaws, Neognathae are the new jaws), and now several strong molecular studies. Some molecular work (Harlid and Arnason 1999) suggested ratites might actually have been derived within the Neognathae, but their results are suspect because of sampling – they sequenced only a crow, a chicken, an ostrich, and a rhea, and therefore got the erroneous results that conflict with more recent studies with much better sampling.


The one very interesting finding that is noted in the Hackett et al. study is the placement of the tinamou within the ratite radiation. Many former classifications list two sister orders among Paleognathae – Stuthioniformes and Tinamiformes. This classification has been supported by previous work, such as the DNA-DNA hybridization of Sibley and Ahlquist or mtDNA studies by van Tuinen et al. (1998). The two groups are separated by the tinamou’s ability to fly, and the placement of tinamou as derived within the ratite lineages could suggest something so radical as the re-evolution of flight within the flightless ratites. Other work does give similar results, including a phylogeny by Chojnowski et al. (2008) based on several nuclear loci. Placing Tinamiformes inside Stuthioniformes in Hackett et al. appears to be a strong result, supported at every locus and partitioning method.

One does not need to invoke the re-evolution of flight to explain the position of the tinamou within the ratites. It is possible the the flighted ratite ancestors spread out across Gondwanaland, where several lineages evolved in parallel to fill similar nitches – the rheas in South America, the Ostrich in Africa, and the Emu, cassowaries, and kiwis in Australia. Meanwhile the tinamous in South America retained the ancestral flighted capabilities. The evolution of a trait in parallel among related lineages has been shown in another group – the whales and the evolution of a nose to a blowhole. If the fossil record for Paleognathous birds is decent, then tracing the evolutionary paths of these groups could be a very fruitful line of future research.
Galloanserae
The next major clade of birds on the tree is another well-known group. This division breaks the Neognathae into the Galloanserae and the Neoaves. The Galloanserae are so named because it is comprised of two orders of bird very familiar to people: Galliformes, the game birds, including pheasants, grouse, quail, and currasows; and Anseriformes, the waterfowl, including ducks, geese, swans, and a few related groups such as the screamers.



I won’t say much about Galloanserae, as the Hackett et al. study posits no changes to the previously recognized ordinal relationships. These results do add additional, significant evidence to these two orders being a valid group, which has been disputed in the past.
Neoaves
Now we move into the Neoaves, containing the vast majority of living bird species, where the real heart of all avian phylogeny problems lies. The problem in reconstructing this phylogeny lies in the fact that it is a rapid radiation – the many major clades are believed to have arisen in such a short time period as to have very short branches between all of the different lineages, resulting effectively in a polytomy that papers such as Hackett et al. have set about trying to resolve. When this radiation of Neoaves occurred is a well-debated subject, revolving around whether the radiation occurred mostly before the K-T boundary when the dinosaurs were still around, or after. The Hackett et al. phylogeny does not seek to address such questions, however they do a good job of demonstrating how closely the various clades are related.
In this tree, the ratites are purple, the Galloanserae are orange, and the Neoaves are the rest.
The biggest division within Neoaves was recently proposed by Fain and Houde (2004). Based on the results of a single nuclear locus, beta-fibrinogen, they divided Neoaves into the Metaves (near-birds) and the Coronaves (crown-birds). This result was quite an upset, because it took several groups previously related groups and split them between Metaves and Coronaves, creating many examples of convergent evolution to ecological niches. A list of examples of feeding niches and comparison of the convergent groups in Metaves and Coronaves are presented by Fain and Houde (2004):


Hackett et al. join at least one other molecular study (Ericson et al. 2006) in recovering the Metaves/Coronaves split, but they call attention to the big caveat. Hackett et al. took the step of examining the effect of each molecular locus used on the topology of the tree, to determine what kind of signal each has. The only loci supporting the Metaves clade are beta-fibrinogen, originally identified in Fain and Houde (2004) and one other locus, out of 19 examined. The fact that only one or two loci drive this division is suspect, as it is possible for genes to form gene trees that are not concordant with the species tree we are attempting to recover. It is also problematic that mitochondrial studies do not recover the Metaves/Coronaves clades (Morgan-Richards et al. 2008, Brown et al. 2008). Determining whether the Metaves/Coronaves clade is an accurate representation of the avian tree of life will depend on further work.
“Metaves”
Because of the uncertainty remaining about the distinction of Coronaves and Metaves, as well as general poor support for ordinal relationships within Metaves, I’ll treat the overall relationships here as unresolved. Several of the groups within Metaves are small families originally placed in larger groups elsewhere. We know they are distinct from their previous relationships although we’re not quite sure where to put them. The best thing worth noting is the presence of several robust groups within Metaves. Even if Metaves is done away with and their place in the overall tree changes, these individual groups are likely to remain valid.

Phaethontidae
The tropicbirds (Phaethontidae) are one of the small families originally placed elsewhere. Tropicbirds are three similar species of pelagic (ocean-going) birds somewhat similar in appearance to birds like gannets (Sulidae), with which they were formerly grouped in Pelecaniformes.

There is not total agreement among the molecular data now available, but Hackett et al. and other studies (Morgan-Richards et al. 2008) suggest that Phaethontidae represents an independent evolution of the pelagic lifestyle in birds. This is at odds with the morphological studies and a few molecular studies (van Tuinen et al. 2001, Brown et al. 2008), which place the tropicbirds with the other totipalmate Pelecaniformes. With morphological studies, there is always the risk that the characters used to build the phylogeny are the ‘wrong’ or uninformative characters. I wonder whether this may be a case of focusing on features more related to feeding morphology that could be arrived upon by convergence on similar lifestyles and other characters would be more informative of the tree the molecular work gives us. I hope more work is done to resolve the position of this family.
Pteroclidae + Columbidae - not a group
Another group split from its former close relatives in Hackett et al. are the sandgrouse (Pteroclidae). This is a group of 16 Old World species that inhabit plains and arid regions. They look like a blend of a dove and a grouse, and they have previously been allied with both of those groups, although almost all evidence now supports a close relationship to the pigeons and doves (Columbidae) with which they are often considered as the order Columbiformes.


Morphology supports a sister relationship between pigeons and sandgrouse (Livezey and Zusi 2007), as do some molecular studies (Ericson et al. 2006). Hackett et al. show the Pteroclidae to have a close relationship with Columbidae (but not sister), but that relationship is one of the poorly supported nodes in Metaves. The precise relationship of Pteroclidae thus continues to remain enigmatic.
Mesitornithidae
A third small family of birds of enigmatic relationship in Metaves are the mesites (Mesitornithidae). This is a group of three terrestrial species endemic to Madagascar that have eluded convincing taxonomic placement. I believe they’ve often been considered in the order Gruiformes (with rails, cranes, and other terrestrial birds), but morphological studies have allied them with the buttonquail (Turnicidae) in the Charadriiformes (Livezey and Zusi 2007), and molecular studies have put them in various relationships. Hackett et al. show some support for a relationship within Metaves with Columbidae. While this relationship has more support than some of the other groups in “Metaves”, it is still relatively weak and it may end up better allied with other groups in Metaves. This is one odd group that deserves focused attention to figure out its proper relationship.

Podicipediformes + Phoenicopteriformes
One well-supported clade in Hackett et al. is the sister relationship of grebes (Podicipediformes) and flamingos (Phoenicopteriformes). Contrary to the reporting in the popular media, this clade was no novel surprise in this study, but was a confirmation of a relatively new phylogenetic hypothesis. The ~20 species of grebe are aquatic diving specialists, with lobed feet and legs positioned so far back on their body for underwater propulsion that they can barely walk. The grebes were originally allied with the loons (Gaviiformes), aquatic divers with similar morphology, but this has long been recognized as a case of convergent evolution on a highly specialized niche, and no recent molecular data support a close relationship of grebes and loons. The flamingos include six living species of charismatic, pink, long-legged, wading, filter-feeding, crazy-billed birds that basically look nothing like grebes. Flamingo relationships have variously jumped around, but the most consistent relationships have been with similar long-legged wading species such as storks and ibises in Ciconiiformes. Some also propose a relationship with the wading birds in the shorebird order Charadriiformes. The grebes and flamingos are two enigmatic taxa with a decent fossil record, with records of modern-appearing grebes and fossils stretching back to the Oligocene (~25 million years ago) but little record before then.

Many molecular studies, regardless of support for Metaves or not, support a sister relationship of grebes and flamingos (van Tuinen et al. 2001, Chubb 2004, Ericson et al. 2006, Brown et al. 2008, Hackett et al. 2008, Morgan-Richards et al. 2008) that I don’t believe had previously been considered. While some morphological phylogenies still cluster the grebes with the convergent loons (Livezey and Zusi 2007), the molecular work prompted Gerald Mayr and others to take a closer look at morphology and fossil evidence. Mayr (2004) found a variety of evidence for morphological synapomorphies (shared derived characters, in other words, unique characteristics that define clades and are not all present in their ancestors) defining a clade of grebes and flamingos. While some have disputed these characters (Storer 2006), virtually all molecular work now supports this relationship so they are probably valid. They include skeletal and muscular characters, including the distinctive shape of certain vertebrae and unique muscle attachment sites on the humerus. Another unique character of this clade is a chalky layer of amorphous calcium phosphate on the eggs, something shared only by Megapodes (Galliformes). A few various other bird groups can have amorphous calcium carbonate layers on the eggs. I don’t know the difference, but it sounds like an interesting feature of the eggs of grebes and flamingos that links them. Overall, the synapomorphies linking flamingos and grebes are potentially difficult to explain as convergences, because of the different lifestyles and corresponding adaptations of the two. Shared recent ancestry seems more likely.
Flamingo foot (Source)

Grebe foot (Source)

Finally, Mayr (2004) notes another morphological character in a fossil flamingo relative that links grebes and loons. Grebes have skeletal canals encasing tendons on the tarsi. In flamingos, the canal is not enclosed, but in the fossil Paleolodus, the canal is partially enclosed. Paleolodus is a fossil genus belonging to a family considered sister to living flamingos, and it is believed to have a more swimming aquatic lifestyle than flamingos, representing an intermediate niche linking what was likely a fully aquatic ancestor to grebes, Paleolodus, and flamingos. Despite this, flamingos and grebes still have very different external foot appearances (see above) which have caused taxonomic confusion before.
All of this evidence combined convinced Mayr to name the flamingo-grebe clade one of the best supported order-level clades in Neoaves. Sangster (2005a) christened the flamingo-grebe clade as the superorder Mirandornithes, the wonderful birds. They truly are.
Eurypygidae + Rhynochetidae
Eurypygidae: Erypygia helias - Sunbittern (Source)

Rhynochetidae: Rhynochetos jubatus - Kagu (Source)

Once more, we have two very different-looking oddball taxa that the molecular work strongly supports a sister relationship for. In this case, we have the monotypic Kagu (Rhynochetidae) of New Caledonia (northeast of Australia), and the monotypic Sunbittern (Eurypygidae) of the Neotropics. They are very different from one another in appearance, although they are similar in habitat: both are terrestrial species of forests. They share powder down, modified feathers that produce a dust they coat their plumage with during preening. They also share a unique behavior of raised wing displays, with contrasting coloration.
Sunbittern wing display (Source)

Kagu wing display (Source)

The sister relationship of these two taxa is not a new proposal. Morphology studies have sometimes placed them as sister taxa within Gruiformes, along with cranes, rails, and finfoots, among others (Cracraft 1982, Livezey and Zusi 2007). Many molecular studies do not sample widely enough to include these two oddball taxa, although several recent studies all support a sister relationship (Fain and Houde 2004, Ericson et al. 2006, Hackett et al. 2008). Fain and Houde (2004) roughly estimate a divergence time of 35 million years ago (Oligocene) around the time of the break-up of the widespread tropical forests that existed at the time. Fain and Houde also cite an Eocene fossil Messelornis as a sister genus to the Sunbittern, and Cracraft (1982) suggest that fossils from New Zealand of the flightless, Moa-like Adzebills belong in Rhynochetidae (although these fossil’s relationships are not solidly resolved), suggesting a much broader distribution from the family. It is likely that the Kagu and Sunbittern are relicts of a lineage after the Gondwanaland breakup. Even if their biogeographic origin and wildly different plumage isn’t well explained, both morphology and molecular data support this puzzling relationship.
Caprimulgiformes + Apodiformes
The final good clade among the ‘Metaves’ that has people scratching their heads indicates that the hummingbirds and swifts (Apodiformes) have evolved within the nightjars and relatives (Caprimulgiformes). The more I consider this proposed clade, the more it makes a lot of sense. In the Apodiformes, we have three families: the hummingbirds (Trochilidae), extremely adapted for aerial, hovering flight to support their nectivory; the treeswifts (Hemiprocnidae) which are long-winged aerial insectivores and quite similar to the swifts (Apodidae), although the treeswifts retain a hind toe that allows them to perch, while the swifts have reversible hind toes that allow them to place all four toes forward (pamprodactyl) and cling to vertical surfaces instead of perching (which they essentially never do). Hummingbirds contain over 300 living species restricted to the New World, with most of their diversity concentrated in the Neotropics. Swifts are found worldwide, with around 80 species, and the treeswifts are four species restricted to southeast Asia.

Caprimulgiformes consists of five to seven families (depending on your splitting preferences). All are cryptically colored, nocturnal birds that feed primarily on insects (with one prominent exception). Many are aerial insectivores and have fairly long wings supporting this lifestyle. The nightjars take this to the extreme, approaching swifts in shape with long, narrow, pointed wings and highly aerial lives. The different families include:
The Oilbird (Steatornithidae), a strange monotypic species in northern South America that is a nocturnal, cave-dwelling forager of palm fruits. This is the oddball taxa of the Caprimulgiformes.

The potoos (Nyctibiidae), another neotropical group of seven species. These birds look and act like branch snags, and are sally out for insect prey at night.
The frogmouths (Podargidae, sometimes split into Australian Podargidae and Asian Batrachostomidae) are 12 species of big, bulky Caprimulgiformes from southeast Asia and Australia.
The nightjars and nighthawks (Caprimulgidae, with eared nightjars sometimes split as Eurostopodidae) are nocturnal aerial insectivores with around 80 species found worldwide.

The owlet-nightjars (Aegothelidae) are 8 Austra\lasia species of long-winged and long-tailed aerial insectivores with really funny-looking faces.

The owlet-nightjars are actually the ‘bridge’ group between the rest of the Caprimulgiformes and the Apodiformes. In Hackett et al., they are found to be the sister group of the Apodiformes, and these families are clustered within the five main Caprimulgiformes lineages. This basically posits that the diurnal super-specialized aerial swifts and the diurnal nectivorous aerial hummingbirds derived from ancestors much like the various lineages of nocturnal aerial insectivores. The alternative hypothesis would be for nocturnal aerial insectivory to have evolved independently among the five main Caprimulgiformes lineages.
This molecular result is not brand-new, but it does lend heavy support for this hypothesis among other hypotheses of Apodiformes and Caprimulgiformes relationships. Not many previous molecular phylogenies sampled enough of the Caprimulgiform lineages to accurate test this hypothesis, but of those that do, Ericson et al. (2006) and Mayr and Clarke (2003) found similar results. A few phylogenies have suggested that Caprimulgiformes and Apodiformes may not be a valid grouping, but most now do, with Livezey and Zusi (2007) and Mayr and Clarke (2003) recovering this relationship in morphological studies. Livezey and Zusi (2007) and Brown et al. (2008) also recover a monophyletic Apodiformes + Caprimulgiformes grouping, but place the Aegothelidae and Apodiformes as a clade sister to the Caprimulgiformes rather than as derived within it. Taken all together, this lends very strong support for Caprimulgiformes and Apodiformes being a valid grouping, and this was recognized as early as 1867 by TH Huxley who named this clade Cypselomorphae. Sangster (2005b) gave the name Daedalornithes to the Aegothelidae + Apodiformes clade, named after Daedalus of Greek mythology who built wings and flew with Icarus. Sangster gave this clade this name to recognize their great flight talents.
 The linking of Apodiformes with the owlet-nightjars and Caprimulgiformes isn’t just the DNA talking. There are actually a growing number of fossils in the Apodiformes lineage that blurs the gaps. In particular, Mayr provides an overview of recently described hummingbird fossils from Europe (!) that show varying stages of development towards the hovering flight style (see his review here). One fossil beats all the rest in suggesting the linkage between the hummingbirds and owlet-nightjars: Parargornis. This ~47 million year old bird is identified as a very early hummingbird based on skeletal characters, yet it has a bill and feathering that resembles an owlet-nightjar (and not a swift).
The linking of Apodiformes with the owlet-nightjars and Caprimulgiformes isn’t just the DNA talking. There are actually a growing number of fossils in the Apodiformes lineage that blurs the gaps. In particular, Mayr provides an overview of recently described hummingbird fossils from Europe (!) that show varying stages of development towards the hovering flight style (see his review here). One fossil beats all the rest in suggesting the linkage between the hummingbirds and owlet-nightjars: Parargornis. This ~47 million year old bird is identified as a very early hummingbird based on skeletal characters, yet it has a bill and feathering that resembles an owlet-nightjar (and not a swift).
One final linking of hummingbirds, swifts, and Caprimulgiformes are certain behavioral traits that pop up in certain lineages. One behavior is torpor – a deep metabolic shutdown that occurs when birds have to deal with extreme demands for energy while keeping warm at night. Torpor is a fairly rare phenomenon in birds, being recorded in only a few families. Interesting, four of those families are swifts, hummingbirds, nightjars, and frogmouths! The only non-Cypselomorphae examples I could find are the swallows and the mousebirds (Coliiformes).
A second behavior linkage may surprise you – echolocation. The oilbird and a clade of swifts known as the swiftlets (Aerodramus and Collocalia) have both evolved the use of audible clicks to help them navigate in dark caves. I don’t know if these behavior traits have been studied to see if they are actual unique features of Cypselomorphae or just independent evolutions of a trait for a particular niche, but it is interesting to note.
As in the flamingo and grebe clade, recent fossil and morphological evidence is supporting new recent molecular hypotheses in the relationships of these birds. Even though the hypothesis of a close relationship of Caprimulgiformes and Apodiformes is nothing new, we now have a strong body of evidence to support it and to show some of the evolutionary history of the group.
“Coronaves”
I’ve covered all of the well-resolved relationships with the purported “Metaves” clade. All of those relationships don’t make up the bulk of avian diversity, however. The bulk lies in the remaining groups of bird, called “Coronaves”, and consists of several well-defined big groups of birds. The relationships among these groups, and whether they all belong as one overall clade “Coronaves” is not resolved with certainty yet, just as is the case with “Metaves” above. The reason for this is once again beta-fibrinogen, rearing its ugly head. Gene-jacknifing analyses in Hackett et al. identify beta-fibrinogen as the main source of information driving the Metaves-Coronaves split. The same gene-jacknifing analyses also identify beta-figrinogen as the sole locus responsible for several relationships within “Coronaves”: the sister relationship between the ‘landbird’ clade and the shorebirds (Charadriiformes) for one, and it also holds together the “waterbirds + core Gruiformes + Turacos + Cuckoos” clade. Once again, I don’t trust the reliance of one locus as a true phylogeny, and unfortunately Hackett et al. do not provide alternative trees that are produced without beta-fibrinogen. In addition, while Hackett et al. resolve many of the inter-clade relationships within “Coronaves”, as opposed to Ericson which left them as several large polytomies, the support for some of these nodes is pretty weak. So, when you collapse the weak nodes of the tree, you get:
Also compounding any review of these groups is the fact that many previous studies recover different results. While I think that the Hackett et al. study is the best phylogeny yet produced, based on the extent of sampling and the number of loci they use in analysis, there is not as much extensive literature to back up some of these clades. Here is an overview of the well-resolved clades that remain.
Waterbirds, Gruiformes, Cuckoos, and more
This clade consists of a large group containing several waterbird orders, the turacos (Musiphagiformes), a third group containing the remaining ‘core’ Gruiformes (i.e. not those split out into “Metaves”) and the cuckoos. As I’ve just stated, the relationships of these four groups – waterbirds, turacos, gruiforms, and cuckoos, are held together in the tree by beta-fibrinogen, and will likely end up not in the between-clade relationships defined in Hackett et al.
 Cuckoos, Turacos, and the Hoatzin - not a group
Cuckoos, Turacos, and the Hoatzin - not a groupOne thing that you can take away from this paper is that the turacos (Musiphagiformes), the cuckoos (Cuculiformes), and the Hoatzin (Opisthocomidae) are likely not closely related. Various previous taxonomies have either kept these as separate, order-level groups, or has lumped them into one, Cuculiformes. I have generally considered keeping these families as one order as a relationship that just kind of ‘makes sense’ based on overall impression of the birds. Indeed, some can approach each other in general shape and habits and they show a progression of foot morphology (Hoatzin are anisodactyl (the default foot morphology), turacos are semi-zygodactyl, cuckoos are zygodactyl).

On the other hand, some of them can look pretty divergent, and cuckoos are mainly insectivorous while turacos are mainly fruit-eaters and the Hoatzin is a leaf-eater. While some morphology studies link them (Livezey and Zusi 2007), no recent molecular phylogeny that I’ve looked at supports Musophagidae and Cuculidae as being sister taxa (Brown et al. 2008, Ericson et al. 2006, Chojnowski et al. 2008). So much for my general impression!



Then there is the Hoatzin. This bird is considered one of the most enigmatic of modern Aves, with no clear relationships emerging even with the molecular phylogenetics of the past 20 years. It has been allied by early molecular data with the cuckoos and turacos, but more recent work has erased that link. Hackett et al. do not solve this puzzle. They place the Hoatzin as a basal member of the waterbird-cuckoo-gruiform clade, but with very weak support. As Christopher Taylor puts it:
The hoatzin is famed for its vile odour and loud call, as it laughs derisively and farts insolently at the unfortunate systematists it has reduced to tears through its intransigence.
So, while the cuckoos seem to cluster pretty well with the Gruiformes, the placement of turacos and the Hoatzin in the Hackett et al. tree is not altogether robust. It is interesting to note that they are all grouping within the same general clade on the tree, even if the exact relationships are not yet known with good support.
core Gruiformes and Bustards
At least four families formerly considered to be in the order Gruiformes have fallen well outside of the other, “core” Gruiformes in the tree in Hackett et al. (Rhynochetidae and Eurypygidae, Seriamidae, Mesitornithidae). Some have referred to this order as a ‘grab-bag’ or ‘trash’ taxon, where lots of families are grouped without very strong support for their actually being related. Well, that certainly appears to be the case. Yet one more split is evident when you examine the overall Gruiformes + Cuculiformes clade – the bustards (Otididae) fall outside of the core Gruiformes with respect to the cuckoos, and indeed the core Gruiformes, the cuckoos, and the bustards are three groups held together in the tree only by beta-fibrinogen, so its possible that the bustards are even farther away from the core Gruiformes than this. The bustards are an Old World family of 25 species. They are big, bulky, terrestrial birds that bring to mind a blend of cranes, grouse, and ratites. They include the heaviest flying bird species (Kori Bustard). Their relationship with the Gruiformes has been based on morphology (Livezey and Zusi 2007), but interestingly, their sister family in that study also has turned out not to be a Gruiform (Cariamidae, see below).
So what’s left? The core Gruiformes is a solid clade consisting of two lineages: In one lineage (Grui): the trumpeters (Psophiidae), 3 species of neotropical terrestrial birds; the cranes (Gruidae), 15 species of tall birds in the Old World and North America; and the Limpkin (Aramidae), a monotypic wading species of the neotropics.

In the other lineage (Ralli): the coots, rails, and moorhens (Rallidae), a worldwide family of 140 species of mostly wetland species (although they occupy many terrestrial niches), and the Sungrebe and finfoots (Heliornithidae), three species of unusual waterbirds found in the Neotropics, Africa, and southeast Asia. These have the incredible unique habit of being able to carry young in flight in pouches in their armpits – I kid you not! Interestingly, a group of African rails called the flufftails (Sarothrura) are found in Hackett et al. to not be in the Rallidae proper but instead are sister to Heliornithidae. A recent phylogeny of the core Gruiformes (Fain et al. 2007) recovered these same relationships, but unfortunately didn’t sample the flufftails to see if they are meritorious of splitting into a different family – I would bet they are, given the otherwise consensus between the two studies.


Waterbirds clade
The big ‘waterbirds’ clade recovered in Hackett et al. was new and surprising for me. It brings together five orders of birds into one monophyletic clade (jumbling a few of them up a bit) united by their common use of aquatic niches. While the focus elsewhere in this review has been on remarkable convergences based on niche adaptations, it is also remarkable to find so many birds of aquatic niches but separate orders still together in one clade. A summary of what overall waterbird relationships used to look like was published by van Tuinen et al. (2001):

Morphology studies more recent than those summarized in van Tuinen continue to show a clade of most of the waterbirds (Livezey and Zusi 2007). The molecular work, all things considered, has not blown those relationships apart, but has removed a few groups already discussed (grebes, flamingos, tropicbirds, etc) and kept most of the remaining waterbird group intact. Almost all recent molecular work shows similar results to those found in Hackett et al., except where poor sampling and few loci prevent the recovery of any decent results (Brown et al. 2008, Chojnowski et al. 2008, Ericson et al. 2008, van Tuinen et al. 2001, etc). It is especially worth noting that, while the mitochondrial and nuclear phylogenies of the avian tree often differ at many points, the recent mtDNA study confirms a waterbird clade and the relationships within it (Morgan-Richards et al. 2008). Hackett et al., as with every other major group covered so far, brings better sampling and better resolution to the tree, so we can be pretty confident in the relations within it.
So, what are the waterbirds? The basal members of the clade are the loons (Gaviiformes), five species of highly modified foot-propelled divers (similar to grebes) that are distributed across the Northern hemisphere.

The remaining four orders are split into two clades. One clade contains the penguins (Sphenisciformes) and the tubenoses (Procellariiformes). The well-known penguins are highly adapted diving birds, flightless and using their flippers (can you really call them wings anymore?) for underwater propulsion. There are about 20 species distributed in the southern hemisphere… okay, really, do I need to talk about penguins much? Everyone knows a penguin.
Procellariiformes are the pelagic tubenoses –ocean-going, highly aerial birds dispersed across every ocean of the world. There are around 125 species in four families, the albatrosses (Diomedeidae), petrels and shearwaters (Procellariidae), the diving-petrels (Pelecanoididae), and storm-petrels (Hydrobatidae). Interestingly, the two sampled storm-petrel genera (Oceanites and Oceanodroma) are not together in the tree, suggesting there are two distinct families. On the other hand, the diving-petrels are found within the Procellariidae in Ericson et al. (2006), suggesting those two families should be lumped. See Penhallurick and Wink (2004) and Kennedy and Page (2002) for review of the familial systematics within Procellariiformes. In the photos below, note how the shearwater is built for flying over the ocean (long, narrow wings to ride the winds) while the diving-petrel is much stockier with shorter wings - built more for diving, yet both retain the same general appearance.

Procellariidae: Puffinus gravis - Great Shearwater (Source)

Pelecanoididae: Pelecanoides garnotii - Peruvian Diving-Petrel (Source)
Pelecanoididae: Pelecanoides garnotii - Peruvian Diving-Petrel (Source)

The sister relationship of penguins with the Procellariiformes isn’t one that makes a whole lot of sense, but the at least some morphology studies (ie Livezey and Zusi 2007) recover this relationship, so there are probably some skeletal traits not immediately apparent that link the diving penguins with the soaring tubenoses. In addition, Wikipedia has a decent review of penguin fossil record, which stretches back at least to 62 million years ago, indicating just how old the divergence between penguins and tubenoses is.
The second clade of waterbirds contains the Ciconiiformes, characterized by tall wading groups (herons, storks, and the like), and the Pelecaniformes, shorter-legged groups with different foraging methods: the cormorants (Phalacrocoracidae), frigatebirds (Fregatidae), gannets and boobies (Sulidae), anhingas (Anhingidae) and pelecans (Pelecanidae). Within this clade of waterbirds, most of the Pelecaniformes families and most of the Ciconiiformes families form distinct lineages… with two notable exceptions.
Basal to either of these two lineages are the storks (Ciconiidae) – around 20 species found almost worldwide. These are large, tall birds of wetlands and grasslands. They include some of the ugliest birds alive, such as this:

The other major exception are the pelecans (Pelecanidae). These are huge, bulky birds well known for their huge gular pouches for scooping up fish and pigeons:
Instead of being found in the clade of the other Pelecaniformes, with which they share such features as totipalmate feet, the pelicans are found within the Ciconiiformes clade, with the herons (Ardeidae), the Hamerkop (Scopidae), the Shoebill (Balaenicipitidae), and the ibises and spoonbills (Threskiornithidae) – all long-legged waders. It is also worth noting that within this group, two odd, different monotypic families yet again turn out to be sister (like the Kagu and Sunbittern): the Hamerkop and the Shoebill from Africa.


The placement of the pelecans and the storks in the waterbird clade has the amusingly ironic result of each family not being found within the order sharing its name! There are at least several previous molecular studies that find this kind of relationship (Ericson et al. 2006, van Tuinen et al. 2001). I look forward to the contortions the taxonomic authorities will have to go through to sort out this mess.
Shorebird Clade - Charadriiformes
The shorebirds stand alone in their own clade, the Charadriiformes, with no real mix-ups presented in Hackett et al. The shorebirds include the sandpipers (Scolopacidae), plovers (Charadriidae), gulls (Laridae), auks (Alcidae), jacanas (Jacanidae), seedsnipe (Thinocoridae) and several more families, and are a diverse group in general united by there preferences for ‘shore’ or aquatic edge habitats – although they have colonized most habitats everywhere from the Himalayas to the Antarctic. Some reporting on the Hackett et al. study report the grouping of the buttonquail (Turnicidae) in the Charadriiformes is a novel result – this is of course not true. Strong molecular phylogenies of the Charadriiformes have been coming out in the last five or so years (many from Alan Baker’s lab) supporting Turnicidae in the Charadriiformes (Paton and Baker 2006, Ericson 2006, to name a few), and new morphological and fossil evidence also places Turnicidae in Charadriiformes (see Mayr 2007 for a review) rather than its former treatment as either a Gruiform or its own order.
Turnicidae: Turnix suscitator - Barred Buttonquail (Source)

Thinocoridae: Thinocorus orbignyianus - Gray-breasted Seedsnipe (Source)
Thinocoridae: Thinocorus orbignyianus - Gray-breasted Seedsnipe (Source)

Landbird Clade
The last remaining grouping of Neoaves, labeled the “landbird clade”, is a grab-bag of what’s left in the avian tree. Unfortunately, while the overall clade has strong support, many of the relationships within this clade were not very strongly supported, so the most practical thing to do is to collapse the unsupported relationships into a polytomy until we can resolve them better. This leaves two big groups – one containing Passerines and a few other surprising relatives, and another containing kingfishers, woodpeckers, and allies – and a few smaller groups of uncertain exact affinities. I’ll start with these smaller groups.

“Falconiformes”
The order Falconiformes, the birds of prey, used to consist of three big families linked by morphology and habits – the scavenging New World vultures (Cathartidae), the cheetahs of the sky, the falcons (Falconidae), and the hawks, eagles, Old World vultures, and kin (Accipitridae). Hackett et al. and a few previous studies support the removal of Falconidae from this group, meaning this is another case where the family sharing the order name has been placed outside of the rest of that order! This leaves Accipitridae, Cathartidae, Pandionidae (the Osprey), and Sagittariidae (the Secretary-Bird) in an order without a proper name – I’ve seen suggests for Accipitriformes already.
Coliiformes

Molecular and morphological studies have bounced the poor african mousebirds (Coliidae) around from group to group, although most of them ally the mousebirds with some member of the ‘landbird clade’. In Hackett et al., their relationships within the landbird clade are not resolved, as their position in the tree receives very weak support. Someday they will find a home…
Strigiformes
The owls (Strigidae, and the barn owls, Tytonidae) are another stand-alone order, like the Coliiformes, that have been bounced around taxonomically. They share traits related to their raptorial habits with the Falconiformes (the old sense, not the new), but they also share traits with the nocturnal Caprimulgiformes, and there have been morphological and molecular studies supporting both views. The owls definitely seem to belong in the land birds clade, but as with all of the land bird groups the exact relationships aren’t yet clear.
Coraciiformes + Piciformes + Trogoniformes
This clade of birds is made up of several fairly similar groups – the Coraciiformes, the Piciformes, and the Trogoniformes – although the distribution of families within this clade mixes up the definitions of those orders. One feature that united these orders was toe arrangement – the trogons have heterodactyl feet, the piciforms have zygodactyl feet, and the coraciiforms have syndactyl feet.
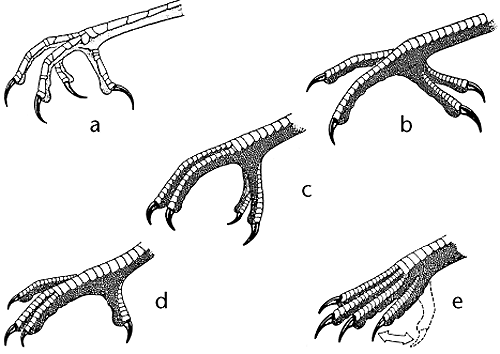
Toe Arrangements (Source) - a) anisodactyl, the default in birds, three forward, one back b) zygodactyl, two forward, two back c) heterodactyl, two forward, two back, but different toes than in zygodactyl d) syndactyl, two forward fused e) pamprodactyl (see swifts)
The Trogoniformes (containing the single family Trogonidae) are cavity-nesters with a unique foot morphology that have been yet another mystery group drifting in their own order.

The Piciformes included the neotropical jacamars (Galbulidae) and puffbirds (Bucconidae), the barbets and toucans (split from one to five families, Megalaimidae, Lybiidae, Capitonidae, Semnornithidae, and Rhamphastidae), the African honeyguides (Indicatoridae), and the woodpeckers (Picidae). In general these groups share a common foot morphology and a burrow- or cavity-nesting tendency.
The Coraciiformes included the kingfishers (Alcedinidae), todies (Todidae), motmots (Momotidae), bee-eaters (Meropidae), rollers (Coraciidae), ground-rollers (Brachypteraciidae), Cuckoo-Roller (Leptosomatidae), Hoopoe (Upupidae), wood-hoopoes (Phoeniculidae), and hornbills (Bucerotidae) (got all that?). The Coraciiformes also share a foot morphology and a common tendency to nest in burrows or cavities.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


Hackett et al. sampled most of these families and found some paraphyletic relationships. The basal member of the overall clade is Leptosomus, the monotypic Cuckoo-Roller from Madagascar. This is a unique bird in appearances, and unlike its former close relatives in Coraciiformes, it actually has semi-zygodactyl feet. The next group in the clade are the trogons. The actual support for their placement here is one of the weaker points in the clade, so it may end up being changed a little, but I think the evidence is now clear that this overall clade is where the trogons finally belong. Then, we come to the Coraciiformes and Piciformes, and discover that the Piciformes is actually nested within two lineages of what we’ve been calling Coraciiformes. The basal lineage of Coraciiformes are the hornbills, hoopoes, and wood-hoopoes of the Old World. The Piciformes maintain their monophyly, with three main groups – the jacamars and puffbirds in the New World, the barbet-toucan clade spread across South America, Africa, and tropical Asia, and the worldwide woodpeckers together with the African honeyguides. The remaining Coraciiformes contain the Old World bee-eaters and roller families, neotropical motmots and todies, and the worldwide kingfishers.


What to do about this taxonomy? This, like the waterbird clade, is going to take a little bit of order re-arranging. I’ve already seen suggestions to name the Hoopoe-Hornbill group the Bucerotiformes or Upupiformes, to retain the traditional core group names of Piciformes and Coraciiformes. I could also see lumping the whole group under one order name, to give attention to the many commonalities throughout the group such as burrow and cavity nesting. On the other hand, the interesting pattern of evolution of foot morphology (and possibly other morphological distinctions that I am sure are present) may warrant recognition.
Seriama-Falcon-Parrot-Passerine clade
This unusual and novel clade was, as best as I have found, first recovered by molecular data in Ericson et al. (2006). The relationships between the four main groups are not all that well worked out – they have poor support on the tree, and were a polytomy in Ericson et al. Mitochondrial data have not shown any support for a clade like this (Morgan-Richards et al. 2009, Brown et al. 2008), and morphology I don’t believe has ever put these groups together. The four groups, briefly, are the Passeriformes, the perching birds, which account for half of the world’s bird species, the parrots, cockatoos, and lories, in the Psittaciformes, the falcons (removed from their former hawk relatives), and the two species of Seriema (Cariamidae), which are odd, long-legged relict species from South America that were formerly allied with the Gruiformes.

There is fairly good support among many recent molecular phylogenies for separating Falconidae from its Falconiformes relatives, although its placement varies (Ericson et al. 2006, Brown et al. 2008, Morgan-Richards et al. 2008, Chojnowski et al. 2008, Fain and Houde 2004). There is less molecular support for separating the seriemas from the Gruiformes, mostly because few phylogenies sample it. Fain and Houde (2004) suggest it is distinct but not in the relationships suggested by Hackett et al.
In general, because this is one of the newest proposals (the closest one of any of those discussed that counts as a ‘new finding’ for the Hackett et al. study), almost no phylogenies recover similar relationships, these are four distinct groups of birds that don’t have distinct relationships with anybody in the tree, and relatively weak support in the tree, I don’t think any changes should be made based on this paper alone. In fact, given these arguments, I am very surprised the this clade is touted as one of the main findings of this paper. I guess you gotta go with what makes headlines…
Final Remarks
Birds are awesome. Figuring out their evolutionary history just makes them even cooler. The new phylogeny in Hackett et al. is certainly not the last, but my intent here was to try to figure out just which groups of birds are almost certainly closely related, and to start to see a window into their evolution.
All throughout this piece, I've stressed the fact that almost nothing in Hackett et al. is a revolutionary hypothesis, despite the popular reporting. In fact, I am skeptical the only truly 'new' hypothesis in this paper, the relationships of seriemas, falcons, parrots, and passerines, precisely because there is no previous body of work to support it (and it is not the strongest relationship in the tree). This paper does not present a total shake-up of the avian tree of life, but it does represent a new landmark for the TOL in completeness and robustness of the tree. Greg Laden has a good discussion on these points, particularly how the modern avian TOL was first really shaken out by the DNA-DNA hybridization of Sibley and Alquist, and everything since then is refinement of the framework that they layed out.
Another major theme throughout the avian TOL is the adapatation to particular niches and particular body plans in many different parts of the tree. Here's just a few examples of the independent evolution of these bird 'types':
Heavy-bodied diving birds - grebes, loons, penguins, diving-petrels, auks, some ducks, and more
Long-winged pelagics - gannets, tubenoses, tropicbirds, some terns
Long-legged waders - flamingos, limpkin, ciconiiformes, some shorebirds
Raptorial birds - hawks, falcons, owls, shrikes
Long-legged savannah birds - ratites, seriemas, secretary-bird
Aerial insectivores - swifts, swallows
Nocturnal predators - owls, frogmouths
These examples don't even fully cover the diversity and many convergences within Passerines! This sort of comparison is just the start of the research possibilities that a robust avian tree of life can lead to. Phylogenies are more than just about systematists keeping themselves busy - they are the pursuit of knowledge and are a necessity for all manner of evolutionary research.
References
Brown, JW, Rest, JS, Garcia-Moreno, J, Sorenson, MD, Mindell, DP. 2008. Strong mitochondrial DNA support for a Cretaceous origin of modern avian lineages. BMC Biology. doi:10.1186/1741-7007-6-6
Chojnowski, JL, Kimball, RT, Braun, EL. 2008. Introns outperform exons in analyses of basal avian phylogeny using clathrin heavy chain genes. Gene, 410(1), 89-96. DOI: 10.1016/j.gene.2007.11.016.
Chubb, AL. 2004. New nuclear evidence for the oldest divergence among neognath birds: the phylogenetic utility of ZENK (i). Molecular Phylogenetics and Evolution, 30(1), 140-151.
Ericson, P.G., Anderson, C.L., Britton, T., Elzanowski, A., Johansson, U.S., Källersjö, M., Ohlson, J.I., Parsons, T.J., Zuccon, D., Mayr, G. (2006). Diversification of Neoaves: integration of molecular sequence data and fossils. Biology Letters, 2(4), 543-547. DOI: 10.1098/rsbl.2006.0523.
Fain, MG, Krajewski, C, Houde, P. 2007. Phylogeny of "core Gruiformes" (Aves: Grues) and resolution of the Limpkin-Sungrebe problem. Molecular Phylogenetics and Evolution, 43:515-529. doi:10.1016/j.ympev.2007.02.015
Fain, MG, Houde, P. 2004. Parallel radiations in the primary clades of birds. Evolution 58(11): 2558-2573.
Hackett, S.J., Kimball, R.T., Reddy, S., Bowie, R.C., Braun, E.L., Braun, M.J., Chojnowski, J.L., Cox, W.A., Han, K., Harshman, J., Huddleston, C.J., Marks, B.D., Miglia, K.J., Moore, W.S., Sheldon, F.H., Steadman, D.W., Witt, C.C., Yuri, T. (2008). A Phylogenomic Study of Birds Reveals Their Evolutionary History. Science, 320(5884), 1763-1768. DOI: 10.1126/science.1157704
Harlid, A, Arnason, U. 1999. Analyses of mitochondrial DNA nest ratite birds within the Neognathae: supporting a neotenous origin of ratite morphological characters. Proceedings: Biological Sciences. 266(1416): 305-309.
Kennedy, M, Page, RDM. 2002. Seabird supertrees: combining partial estimates of procellariiform phylogeny. The Auk. 119(1): 88-108.
Livezey, BC, Zusi, RL. 2001. Higher-order phylogenetics of modern aves based on comparative anatomy. Netherlands Journal of Zoology. 51(2): 179-205.
Livezey BC, Zusi RL. 2007. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zoological Journal of the Linnaean Society. 149: 1-95.
Mayr, G. 2003a. Phylogeny of early Tertiary swifts and hummingbirds (Aves: Apodiformes). The Auk 120(1): 145-151.
Mayr, G. 2003b. A new Eocene swift-like bird with peculiar feathering. Ibis 145: 382-391.
Mayr, G. 2004. Morphological evidence for sister group relationship between flamingos (Aves: Phoenicopteridae) and grebes (Podicipedidae). Zoological Journal of the Linnaean Society. 140: 157-169.
Mayr, G. 2005. Fossil hummingbirds in the Old World. Biologist 52(1): 12-16. (link)
Mayr, G. 2007. The renaissance of avian paleontology and its bearing on the higher-level phylogeny of birds. Journal of Ornithology. 148: S455-S458.
Mayr, G. 2007. Avian higher-level phylogeny: well-supported clades and what we can learn from a phylogenetic analysis of 2954 morphological characters. J Zool Syst Evol Research. 46(1): 63-72. doi: 10.1111/j.1439-0469.2007.00433.x
Mayr, G, Clarke, J. 2003. The deep divergences of neornithine birds: a phylogenetic analysis of morphological characters. Cladistics 19: 527-553. doi:10.1016/j.cladistics.2003.10.003
Mayr, G, Manegold, A, Johansson, US. 2003. Monophyletic groups within 'higher land birds' - comparison of morphological and molecular data. J Zool Syst Evol Research. 41: 233-248.
Mindell, DP, Sorenson, MD, Dimcheff, DE, Hasegawa, M, Ast, JC, Yuri, T. 1999. Interordinal relationships of birds and other reptiles based on whole mitochondrial genomes. Systematic Biology. 48(1): 138-152.
Morgan-Richards, M, Trewick, SA, Bartosch-Harlid, A, Kardailsky, O, Phillips, MJ, McLenachan, PA, Penny, D. 2008. Bird evolution: testing the Metaves clade with six new mitochondrial genomes. BMC Evolutionary Biology, doi:10.1186/1471-2148-8-20
Paton, TA, Baker, AJ. 2006. Sequences from 14 mitochondrial genes provide a well-supported phylogeny of the Charadriiform birds congruent with the nuclear RAG-1 tree. Molecular Phylogenetics and Evolution. 39(3): 657-667. doi:10.1016/j.ympev.2006.01.011
Penhallurick, J, Wink, M. 2004. Analysis of the taxonomy and nomenclature of the Procellariiformes based on complete nucleotide sequences of the mitochondrial cytochrome b gene. Emu 104: 125-147.
Sangster, G. 2005a. A name for the flamingo-grebe clade. Ibis 147: 612-615.
Sangster, G. 2005b. A name for the clade formed by owlet-nightjars, swifts, and hummingbirds (Aves). Zootaxa 799: 1-6.
Storer, RW. 2006. The grebe-flamingo connection: a rebuttal. The Auk. 123(4): 1183-1184.
van Tuinen, M, Sibley, CG, Hedges, SB. 1998. Phylogeny and biogeography of ratite birds inferred from DNA sequences of the mitochondrial ribosomal genes. Mol Biol Evol 15(4): 370-376.
van Tuinen, M, Butvill, DB, Kirsch, JAW, Hedges, SB. 2001. Convergence and divergence in the evolution of aquatic birds. Proceedings: Biological Sciences. 268(1474): 1345-1350
Great post Nick. I was trained as a plant breeder many years ago and I ofter wondered why more work had not been done genetically with birds, especially now that there is all that cool software to run the tests. Again, Good Work.
ReplyDeleteThanks! It only took over a week to research and write :)
ReplyDeleteWell, I'm impressed. There's only a couple of points I'd quibble about.
ReplyDeleteThe sister relationship of penguins with the Procellariiformes isn’t one that makes a whole lot of sense
You know, I've always thought that it makes a great deal of sense. Tubular nostrils are actually found in juvenile little blue penguins (Eudyptula minor) as well as in Procellariiformes, for a start.
While of course not directly relevant to the possibility of a procellariiform-spheniscid relationship, a very similar pattern of relationship occurs in the Northern Hemisphere with the close relationship of the highly volant Laridae and the tubbier Alcidae. The alcids mostly haven't lost flight, of course, having more of a diving habitus [the strong superficial resemblance between diving petrels (Pelecanoides) and little auks (Alle alle) is legendary), but they do include the original "penguin" (which the Southern Hemisphere birds were actually named after), Pinguinus impennis.
Interestingly, a group of African rails called the flufftails (Sarothrura) are found in Hackett et al. to not be in the Rallidae proper but instead are sister to Heliornithidae.
I ovrlooked this! It is notable that flufftails have long been regarded as one of the basalmost groups of rails, with only Himantornis haematopus more basal. To the best of my knowledge, Himantornis has never been sampled in a molecular phylogeny. Someone needs to go on a trip to central Africa to get some genes.
This leaves Accipitridae, Cathartidae, Pandionidae (the Osprey), and Sagittariidae (the Secretary-Bird) in an order without a proper name – I’ve seen suggests for Accipitriformes already.
Accipitriformes as a separate order from Falconiformes for Accipitridae and its closest relatives has been around for some time, and was used in sources even predating Sibley and Ahlquist.
Clark's Grebe
Funnily enough, all the pictures I've seen of Clark's Grebe were from the side - I never realised they had that funky hairdo.
Cuckoo-Roller
The bird with the biggest head ever. Mein Gott, it looks like a child's doodle-bird.
I’ve already seen suggestions to name the Hoopoe-Hornbill group the Bucerotiformes or Upupiformes
Again, been about for a while. IIRC, Bucerotiformes would be the entire group, or the hornbills alone, while Upupiformes would be hoopoes if regarded as a separate order from the hornbills.
Seriama-Falcon-Parrot-Passerine clade
Morphological data has been used to support a relationship between seriemas and falcons, though maybe not exclusively. Darren Naish wrote on it a couple of years ago.
Thanks, man! Your comments are the best.
ReplyDeletePenguins - I didn't know about the tubular nostrils in those penguins. I'll have to look into that further.
Flufftails - look at this response:
http://www.birdforum.net/showthread.php?t=117864
It suggest Himantornis isn't basal, but those results aren't published to my knowledge.
Order names - I didn't realize those names were all previously proposed. That would explain why people are so quick to pick them up.
Thanks for the link to Darren's work. I've been thinking about putting some extinct birds into this work (moas, which we have molecular sequences for, and phorusrhacids, because they are cool) but I didn't get to it.
~ Nick
Just wanted to add my thanks for your efforts on this; it was very interesting.
ReplyDeleteThank you, for reading! :)
ReplyDeleteI can not believe that cuckoos and turacos are not related to owls and parrots. They have all zygodactyl feet, what about this?
ReplyDeleteHi there, Ive only just found your blog!? Been reading through your posts past to present and found some really interesting stuff! This particular article interests me as I'm currently writing my dissertation looking at Avian taxonomy and its transition from morphological to molecular phylogeny. I also greatly enjoyed reading your articles titled 10,000 birds? part one and two. Amazing.
ReplyDeleteBeautiful Birds!
ReplyDelete